 |
|||||||||||||||
A célula como unidade da Vida |
É um dos pilares fundamentais da Biologia o facto de que todos os organismos vivos săo formados por células, e que todas as células provęm de uma célula preexistente. Estas afirmaçőes constituem, basicamente, a teoria celular. Esta teoria tem algumas implicaçőes importantes, nomeadamente que o conhecimento da biologia celular é fundamental para o conhecimento da Vida. Revela, igualmente, que a Vida é contínua, pois o nosso corpo formou-se a partir de uma célula (zigoto) e irá produzir outras células (óvulos e espermatozóides) que, por sua vez, formarăo novos corpos, e assim sucessivamente. As células săo sempre muito pequenas, sendo necessário um microscópio para as observar correctamente. As honrosas excepçőes săo os óvulos de muitas espécies, com os das aves em grande destaque. Este facto deve-se ŕ importância da relaçăo área/volume, que vai sendo cada vez menor com o aumento de tamanho da célula.
|
||||||||||||||
| Célula procariótica |
As células procarióticas săo características dos organismos dos Domínios Bacteria e Archea, razăo porque estes săo designados procariontes. Os organismos procariontes săo unicelulares mas muitos deles formam colónias de forma linear ou em pequenos grupos. As células procarióticas săo geralmente menores que as eucarióticas, variando entre 0,25x1,2 um e 1,5x4 um. Por este motivo, embora visíveis ao M.O.C., a sua substrutura apenas pode ser estudada ao M.E. Este tipo de célula năo apresenta compartimentos membranares no seu interior, ao contrário das células eucarióticas, estudadas a seguir. Apesar de serem estruturalmente mais simples que as eucarióticas, as células procarióticas săo funcionalmente complexas, sendo capazes de realizar milhares de transformaçőes bioquímicas.
Todas as células eucarióticas apresentam as mesmas características básicas:
Algumas bactérias săo fotossintéticas (cianobactérias) apresentam pregas e dobras da membrana citoplasmática que formam um sistema membranar - lamelas fotossintéticas - contendo bacterio-clorofila e outros pigmentos necessários ŕ captaçăo da energia luminosa. Outros grupos de procariontes apresentam outro tipo de estruturas membranosas designadas mesossomas, cuja funçăo pode estar relacionada com a divisăo celular ou com a produçăo de energia. Tal como as lamelas fotossintéticas, os mesossomas săo dobras da membrana citoplasmática e nunca dela se libertam (năo săo organitos livres no citoplasma, como os das células eucarióticas). Por vezes as bactérias deslocam-se com a ajuda de apęndices designados flagelos. Estes apęndices săo formados por uma proteína - flagelina - e geralmente parecem pequenos saca-rolhas ao M.O.C. Os flagelos săo extremamente eficientes na deslocaçăo das bactérias, rodando (em ambos os sentidos) em volta de anéis proteicos que os ligam ŕ membrana citoplasmática e mesmo ŕ parede, se esta está presente. Com o flagelo a bactéria desloca-se a cerca de 0,00015Km/h, o que pode năo parecer impressionante, a năo ser que se faça a seguinte comparaçăo:
Semelhantes a flagelos mas mais curtos e numerosos (cerca de 3 ou 4 por célula) săo os pili, embora a sua funçăo seja algo diferente, aderęncia e transferęncia sexual. Durante a conjugaçăo, um tipo especial de pilum, o F-pilum, é usado para transferir material genético de uma bactéria dadora para uma bactéria receptora. As fímbrias săo mais curtas e lineares que os flagelos e pili, mas năo tęm qualquer funçăo na mobilidade bacteriana. Geralmente usados para aderir a célula, tanto a outra (durante o acasalamento, por exemplo), como a superfícies ou organismos para obtençăo de alimento e protecçăo.
|
||||||||||||||
| Origem da célula eucariótica |
A
maioria dos biólogos considera que a divisăo fundamental no mundo biológico
é a que separa os seres procariontes dos eucariontes, divisăo esta,
baseada na estrutura celular dos organismos. Os
procariontes constituem, mesmo na actualidade, mais de metade da biomassa da
Terra, e colonizaram todos os ambientes. No entanto, a evoluçăo năo se
satisfez com este sucesso e surgiram níveis mais complexos de organizaçăo. A origem da Vida parece ter ocorrido há cerca de 3400 M.a., quando o nosso planeta já teria 1000 ou 1500 M.a. de idade. A célula conserva em si, a nível da sequęncia de aminoácidos, proteínas ou bases nucleotídicas, diversas marcas do seu passado, pois cada gene de uma célula actual é uma cópia de um gene muito antigo, ainda que com alteraçőes. Este é o motivo porque se considera a existęncia de um ancestral comum entre organismos que apresentem grande número de nucleótidos ou proteínas comuns.
Até
há pouco tempo considerava-se que as células eucarióticas teriam derivado de
procariontes unicelulares, por um
processo desconhecido de complexificaçăo,
designado por hipótese autogénica. Esta teoria
considera que a célula eucariótica teria surgido através de especializaçăo
de membranas internas, derivadas de invaginaçőes da membrana plasmática. É
sabido que a associaçăo entre duas células é comum e pode trazer vantagens
importantes, tanto em procariontes como em eucariontes. Surge, portanto, um corolário para esta afirmaçăo, que consiste na obrigatoriedade da presença de células eucarióticas para o desenvolvimento da multicelularidade. A teoria de maior aceitaçăo, proposta por Lynn Margulis, a Teoria Endossimbiótica, sugere que as células eucarióticas seriam o resultado da associaçăo de células procarióticas simbióticas.
A
simbiose entre estas células procarióticas teria evoluído para graus de
intimidade tais, que algumas células envolveriam outras completamente, embora
as primeiras ficassem intactas no interior do hospedeiro. Estas células
envolvidas teriam originado os organitos de uma célula eucariótica actual. Segundo Margulis, a célula eucariótica típica teria surgido sequencialmente, em 3 etapas, como se pode ver ao lado:
Um bom exemplo de como esta teoria pode ser correcta é a evoluçăo dos cloroplastos em protistas fotossintéticos, que parece resultar de uma série de processos endossimbióticos. Aparentemente todos os cloroplastos remontam ao envolvimento de uma cianobactéria ancestral por uma outra célula, um proto-eucarionte. Este será designado o fenómeno endossimbiótico primário e teria resultado na formaçăo do cloroplasto clássico com duas membranas (uma resultante da membrana plasmática da cianobactéria e outra da membrana da vesícula de endocitose da célula maior). Teria sido assim que surgiram os cloroplastos das algas verdes e vermelhas. As algas euglenófitas, no entanto, teriam cloroplastos formados por um fenómeno endossimbiótico secundário, ou seja, o seu ancestral terá envolvido uma clorófita unicelular e descartado toda a célula excepto o cloroplasto. Esta é uma possível explicaçăo para o facto de as euglenófitas apresentarem os mesmos pigmentos fotossintéticos que as clorófitas e as plantas, bem como para a terceira membrana que envolve o cloroplasto destas algas unicelulares. Outros protistas fotossintéticos apresentam cloroplastos resultantes da endossimbiose secundária de rodófitas unicelulares e chegam mesmo a participar em fenómenos de endossimbiose terciária, originando um grupo de dinoflagelados com cloroplastos envolvidos por quatro membranas. Alguns outros
factos parecem apoiar
a teoria endossimbiótica:
No
entanto, muitas dúvidas persistem, pois a transferęncia lateral de genes
complica grandemente o estudo das linhagens celulares mas, ao mesmo tempo, năo
parece ter sido suficiente para explicar o facto de cada vez mais genes com
origem bacteriana serem encontrados em eucariontes. Uma
origem endossimbiótica de mitocôndrias e cloroplastos permite explicar a
presença de genes bacterianos que codificam enzimas do metabolismo energético
mas năo explica a presença de muitos outros. O genoma eucarionte é claramente
uma mistura com dupla origem. Um sugestăo recente propőe que o domínio Eukarya tenha surgido através de uma fusăo mutualista (e năo uma endossimbiose) de uma bactéria Gram – e de uma arqueobactéria, mas ainda precisa de mais provas.
|
||||||||||||||
| Célula eucariótica |
Além do núcleo, as células eucarióticas apresentam uma grande variedade de organitos, ausentes nos procariontes, nomeadamente compartimentos membranares com ambientes físico-químicos diferentes do citosol, o que permite a realizaçăo de reacçőes bioquímicas específicas. Outra característica única das células eucarióticas é a presença de citosqueleto, que lhes fornece suporte e mecanismos para o movimento. O núcleo é geralmente o maior organito da célula, funcionando como centro de controlo da célula e como local onde decorrem a armazenagem e replicaçăo do DNA. Em células metabolicamente activas é frequente observar um ou mais nucléolos, locais onde se formam os ribossomas, a partir de proteínas específicas e RNA. Os nucléolos năo estăo isolados por membranas do resto do núcleo. O núcleo está envolvido por uma dupla membrana designada invólucro nuclear. Este invólucro é atravessado por numerosos poros com dimensőes entre os 3 e os 100nm, que estabelecem a comunicaçăo entre o nucleoplasma e o citosol. Estes poros năo săo apenas buracos mas tęm uma estrutura proteica complexa: cada poro é rodeado por 8 proteínas em forma de grânulo. Nestas zonas a membrana externa do núcleo é contínua com a membrana interna. A membrana nuclear está frequentemente em contínuo com o retículo endoplasmático. O interior do compartimento nuclear - nucleoplasma - é uma soluçăo aquosa em tudo semelhante ao citosol, que contém uma rede de filamentos associados ao invólucro nuclear e ŕ cromatina. A cromatina é um complexo de DNA e proteínas designadas histonas, organizadas em longos filamentos durante a maior parte do ciclo celular. Apenas durante a divisăo celular estes filamentos se condensam, formando cromossomas, visíveis ao M.O.C. A membrana citoplasmática é responsável pela integridade da célula, bem como pela regulaçăo da passagem de moléculas para o interior e/ou para o exterior. É formada por uma bicamada fosfolípidica onde se encontram embebidas proteínas e outras moléculas (colesterol, por exemplo, em células animais). A membrana envolve todo o compartimento celular, que contém o citosol e os organitos. O citoplasma das células está frequentemente em movimento, arrastando de modo ordenado os organitos e os materiais em suspensăo. Estes movimentos, conhecidos como correntes citoplasmáticas ou movimentos de ciclose, devem favorecer as trocas entre os diversos componentes celulares mas năo é certo que seja essa a sua funçăo principal. O retículo endoplasmático é parte do extenso sistema endomembranar que compőe a maioria do citoplasma de uma célula eucariótica. Em certas locais é contínuo com o invólucro nuclear. O retículo pode ser funcional e estruturalmente dividido em retículo endoplasmático rugoso e liso. O retículo rugoso deve o seu nome ao facto de as suas membranas conterem ribossomas, locais de síntese proteica. As proteínas săo lançadas para o interior das membranas, onde serăo transformadas ou dirigidas a outras localizaçőes da célula. O retículo liso năo apresenta ribossomas nas suas membranas e nele as proteínas sintetizadas no retículo rugoso săo quimicamente alteradas. Ocorrem ainda no seu lúmen a hidrólise do glicogénio, síntese de esteróides e alteraçăo de drogas e outras substâncias nocivas ao corpo. O aparelho de Golgi deve o seu nome ao cientista italiano que primeiro o observou ao microscópio óptico composto. É formado por sáculos membranosos achatados designados cisternas e pequenas vesículas. O aparelho de Golgi recebe do retículo endoplasmático rugoso (R.E.R.)proteínas, transportadas em vesículas membranosas. Essas moléculas săo entăo separadas e modificadas quimicamente, sendo depois encaminhadas para as suas localizaçőes definitivas. Se se destinam ao exterior da célula săo "embaladas" em vesículas que se irăo fundir com a membrana plasmática e libertadas para o exterior. Se se destinam ao citoplasma, as vesículas irăo fundir-se com outros organitos. As vesículas formadas pelo R.E.R. săo recebidas pela face cis, ou de recepçăo, do Golgi, as proteínas alteradas no lúmen das cisternas e libertadas pela face trans, ou de formaçăo, virada para a membrana plasmática. A mitocôndria é o organito responsável pela transformaçăo da energia contida nos alimentos em energia metabólica (ATP). Estas transformaçőes designam-se, no seu conjunto, respiraçăo celular. As mitocôndrias tęm duas membranas, como o núcleo ou os cloroplastos. A membrana externa é lisa e fornece protecçăo, embora seja bastante permeável á passagem de substâncias. A membrana interna contém grandes complexos proteicos embebidos, envolvidos na síntese de ATP e na respiraçăo celular. Esta membrana está dobrada em pregas achatadas designadas cristas, que aumentam grandemente a sua área. O número de cristas varia muito com a taxa metabólica da célula em que a mitocôndria se encontra. A matriz é a regiăo interna da mitocôndria, rodeada pela membrana interna. Contém numerosas proteínas envolvidas nos processos respiratórios, bem como ribossomas e DNA, usados na síntese da maioria das suas proteínas. O peroxissoma é um organito relativamente pouco conhecido, em que produtos tóxicos para a célula, como peróxido de hidrogénio (água oxigenada), săo degradados em produtos inofensivos, como água e oxigénio. Exclusivamente em células vegetais existem organitos semelhantes, designados glioxissomas. O centrossoma é uma regiăo mais ou menos amorfa, localizada perto do envelope nuclear em células animais. Ao centro desta zona está um par de estruturas cilíndricas designadas centríolos e dispostas em ângulo recto, como um L. Os centríolos estăo intimamente relacionados com o movimento celular, seja por meio de flagelos ou de cílios, em cuja base existe sempre um corpo basal, em tudo semelhante ao centríolo típico. Esta relaçăo é confirmada pela facto de muitas vezes os flagelos ou cílios serem reabsorvidos e os seus corpos basais deslocados para o interior da célula, passando a funcionar como centríolos. A estrutura do centríolo é formada por nove grupos de tręs microtúbulos fundidos. Estes nove conjuntos formam a "parede" da estrutura, ligeiramente rodados para o interior, como hélices de uma turbina. Cada conjunto está ligado longitudinalmente ao adjacente por outro tipo de proteínas. Característico das células eucarióticas (e talvez, por ausente das bactérias, um factor fundamental no sucesso dos eucariontes), o citosqueleto é formado por um emaranhado de longas fibras de vários tipos, fornecendo suporte e permitindo o movimento (seja da célula ou de organitos ou cromossomas no seu interior) e a alteraçăo de forma. Devido ao seu elevado dinamismo e interacçăo com o ambiente, poderia, com a mesma facilidade, designar-se citomusculatura. O citosqueleto é responsável pelo deslizar sobre o substrato, contracçăo muscular e pelas alteraçőes de forma durante o desenvolvimento embrionário dos animais. O citosqueleto contém 3 tipos principais de fibras:
Os plastídeos săo geralmente classificados segundo o tipo de pigmentos que contęm. Os cloroplastos săo plastídeos que apenas existem em células autotróficas, ditas vegetais. As células animais, heterotróficas, năo produzem cloroplastos mas podem apresentá-los, perfeitamente funcionais, retirados da digestăo parcial de células vegetais ou devido a algas verdes que vivem em simbiose nesses tecidos. Esta situaçăo é bastante comum em corais e anémonas. É o local onde se realiza a fotossíntese, pelo que contém grande quantidade de pigmentos, nomeadamente clorofilas. Uma única célula do mesófilo pode conter até 50 cloroplastos, pelo que 1mm2 de folha contém cerca de 500000. Geralmente localizam-se nos lados longos das células, perto da parede celular. A sua estrutura faz lembrar a da mitocôndria, pois também apresenta duas membranas. A membrana externa é bastante permeável, permitindo a passagem da maioria das pequenas moléculas. A membrana interna é bem mais selectiva e é onde se localizam os complexos que captam a luz para as reacçőes fotossintéticas. Forma dobras designadas tilacóides. Quando os tilacóides aparecem empilhados como moedas designam-se grana. O espaço interno do cloroplasto designa-se estroma e é rodeado pela membrana interna. Neste espaço, tal como na mitocôndria, encontra-se DNA e ribossomas, capazes de comandar numerosas proteínas presentes no cloroplasto. No entanto, o controlo é nitidamente do núcleo, sendo a maior parte do material sintetizado com DNA nuclear e transferido para o plastídio. Em algas verdes e plantas é frequente encontrar no estroma grăos de amido e/ou pequenas gotas de lípidos. Estes săo produtos de armazenamento temporário, quando o organismo fotossintetiza activamente. Os cromoplastos săo plastídeos de tamanhos muito diversos que armazenam outro tipo de pigmentos (que năo clorofila), principalmente carotenóides. Săo responsáveis pelas cores amarela, laranja e vermelha de folhas velhas, flores e frutos maduros. Podem desenvolver-se a partir de cloroplastos em que a clorofila e as membranas internas se desintegram e grandes quantidades de carotenóides săo armazenados. A sua funçăo na planta năo é bem reconhecida, embora sejam fundamentais na atracçăo de insectos e vertebrados, com os quais as plantas evoluíram. Os leucoplastos săo plastídeos năo pigmentados. Alguns sintetizam amido - amiloplastos - outros prótidos e mesmo lípidos. Quando expostos ŕ luz transformam-se em cloroplastos. A parede celular é uma estrutura semi-rígida presente nas células vegetais externamente ŕ membrana citoplasmática. É basicamente formada por celulose mas contém igualmente polissacáridos complexos e proteínas. A parede permite a manutençăo da forma da célula vegetal, impedindo a sua
lise, é uma barreira bastante eficaz contra ataques de microrganismos
patogénicos e ajuda na uniăo entre células
vizinhas. |
||||||||||||||
| Importância da relaçăo área/volume e origem da multicelularidade |
Há medida que as dimensőes de um organismo aumentam, diminui a sua relaçăo área/volume, ou seja, a sua superfície năo aumenta ŕ mesma taxa que o volume. Qual o significado deste facto ? A vida depende do metabolismo, efectuado em todo o volume celular, mas as trocas com o meio, nomeadamente a entrada de nutrientes e a saída de excreçőes, săo realizadas através da superfície celular.
Este raciocínio permite compreender
facilmente que haverá uma razăo óptima para a qual as trocas săo adequadas
ao metabolismo desenvolvido. Estudos revelaram que esse valor corresponde ao tamanho da célula eucariótica, 50 a 500 mm. A partir deste valor o aumento de tamanho de um organismo implica a passagem ŕ multicelularidade, para que a relaçăo correcta seja mantida. No entanto, mesmo
a multicelularidade apresenta a mesma limitaçăo pois os organismos muito
pequenos perdem demasiado calor, e os grandes tęm grande dificuldade em
perde-lo, por exemplo. A
verdadeira multicelularidade, apenas presente em seres eucariontes,
caracteriza-se por uma associaçăo de células em que há interdependęncia
estrutural e funcional entre elas. Geralmente existe igualmente uma diferenciaçăo
celular e tecidular a ela associada. Em
células eucarióticas existe frequentemente uma relaçăo colonial, que pode
ser considerada a origem da multicelularidade. Vejamos
alguns exemplos, que parecem confirmar essa hipótese:
A
evoluçăo destas colónias terá sido por aumento do número e tamanho das suas
células, aumento da especializaçăo e desenvolvimento progressivo das
estruturas sexuais. A dificuldade reside na passagem de uma colónia esférica para um metazoário, com várias camadas de células. |
||||||||||||||
|
Temas relacionados: |
|||||||||||||||
|
|||||||||||||||


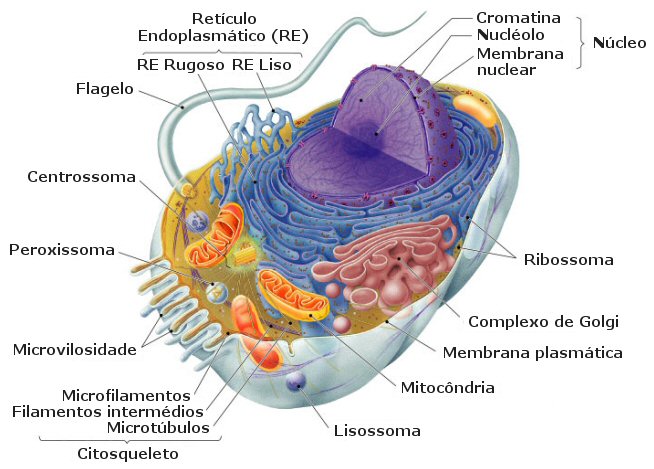 A célula
eucariótica, como a célula animal representada ao lado, obtém a sua designaçăo do grego eu = verdadeiro + karyon
= núcleo. Estas células existem em quase todos os organismos vivos actuais, com
excepçăo dos pertencentes aos Domínios Archea e Bacteria.
A célula
eucariótica, como a célula animal representada ao lado, obtém a sua designaçăo do grego eu = verdadeiro + karyon
= núcleo. Estas células existem em quase todos os organismos vivos actuais, com
excepçăo dos pertencentes aos Domínios Archea e Bacteria.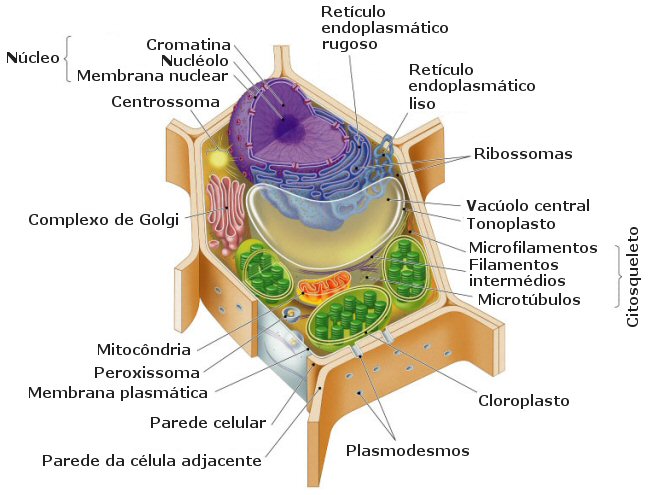 Os
plastídios săo característicos das células
vegetais como a representada ao lado e tęm uma estrutura característica: săo envolvidos por um envelope
com duas membranas, a mais interna das quais se diferencia num sistema
complexo, e que rodeia uma matriz mais ou menos homogénea, o estroma.
Os
plastídios săo característicos das células
vegetais como a representada ao lado e tęm uma estrutura característica: săo envolvidos por um envelope
com duas membranas, a mais interna das quais se diferencia num sistema
complexo, e que rodeia uma matriz mais ou menos homogénea, o estroma. 

